Inspirationsdag 8. januar 2010 Workshop: Bioteknologi fra gen til proteindesign
|
|
|
- Jacob Justesen
- 8 år siden
- Visninger:
Transkript
1 Inspirationsdag 8. januar 2010 Workshop: Bioteknologi fra gen til proteindesign Arrangeret af Sektion for Biomolekylære Videnskaber, Biologisk Institut, Københavns Universitet Ansvarlige: Karen Skriver, lektor, Helle Blæsild, kommunikationsmedarbejder, Birthe B. Kragelund, lektor, Kaare Teilum, adjunkt, Martin Willemoës, lektor, Gert Dandanell, lektor, Michael Riis Hansen, post. doc, Jakob Winther, professor, Januar 2010 [] 1

2 Workshoppen Bioteknologi fra gen til proteindesign giver mulighed for at arbejde eksperimentelt og in silico med enzymet alkalisk fosfatase i forsøg, som vil kunne implementeres i gymnasieundervisningen. Der vil også være mulighed for at diskutere forskningsområder inden for proteinstruktur, -design og - engineering med forskere, der kan bestilles som foredragsholdere i gymnasiet. Program : Intro. (rum ) Bioteknologi fra gen til proteindesign ved Karen Skriver : Programpakke 1 (rum ; 6 deltagere): Produktion og oprensning af alkalisk fosfatase. Ansvarlige: Karen Skriver og Charlotte O Shea. Programpakke 2 (rum ; 12 deltagere): Kinetiske analyser af alkalisk fosfatase. Ansvarlige: Michael Riis Hansen og Gert Dandanell. Programpakke 3 (rum ; 10 deltagere): Strukturanalyser af alkalisk fosfatase. Ansvarlige: Birthe Kragelund og Kaare Teilum. Præsentation af forskningsområder i rum Martin Willemoës Design af enzymer Enzymdesign er en fremtidig teknologi med et enormt potentiale. Enzymer bruges f. eks. i vaskepulver, papirindustri over fødevareindustri til avanceret blødermedicin og i tilvejebringelsen af miljøvenlig biobrændsel. Den begrænsning, som samtidig er en eftertragtet egenskab i anvendelsen af enzymteknologi, er specificiteten overfor et substrat; givet at man har været afhængig af naturligt forekommende enzymer som de hovedsageligt findes i mikroorganismer. Ved i heldige tilfælde at ændre lidt på enzymets egenskaber har man kunnet udvide det "rum" af enzymaktiviteter som naturen har stillet til rådighed. Forskningsgruppen Protein Design Consortium arbejder langsigtet på at opstille metoder, både eksperimentelle og computermæssige, der skal muliggøre frit at kunne designe et enzym, der kan katalysere en hvilken som helst kemisk reaktion. Dette arbejde og vores udgangspunkt kan du høre nærmere om. Kaare Teilum Protein engineering som værktøj til at forstå sygdomsrelateret proteinudfældning I en række alvorlige sygdomme såsom type II diabetes, Alzheimers- og Parkinsons sygdom samt amyotrofisk lateral sklerose (ALS) udfælder specifikke protein i de berørte væv. Udfældningerne har ofte karakter af fibriller (små fibre af protein). På trods af den tydelige ophobning af fibriller i disse sygdomme tyder meget på, at fibrillerne i sig selv, ikke er farlige for det væv, de befinder sig i. Det, der giver anledning til sygdommene, er derimod snarere mindre forstadier til fibrillerne, og der har gennem de seneste år været stor interesse for at forstå, hvordan og hvorfor fibrilforstadierne giver anledning til sygdomme. En vigtig brik i denne forståelse ville være at kende strukturerne af fibrilforstadierne, men det er indtil videre ikke lykkedes at bestemme disse. Ved hjælp af protein engineering kan forskellige forstadier i udfældningen af de enkelte proteiner stabiliseres i tilstrækkelig grad til, at deres strukturelle egenskaber kan analyseres. Her kan du høre om, hvordan [] 2
 Bioteknologi fra gen til proteindesign ved Karen Skriver 14.40 17.00: Programpakke 1 (rum 2-0-14; 6 deltagere): Produktion og oprensning af alkalisk fosfatase.")
3 protein engineering bruges i klarlæggelsen af, hvordan proteinet superoxid dismutase danner udfældninger i ALS. Jakob Winther Forudsigelse af proteinstrukturer fra aminosyresekvenser og omvendt Proteiners struktur defineres af rækkefølgen af de aminosyrer som de er opbygget af. Aminosyrekæden folder i proteiner op i mere eller mindre komplicerede tredimensionelle strukturer (som fx vist på forsiden af denne folder) og proteiners funktion vil almindelighed være fuldstændig afhængige af at strukturen dannes rigtigt. Dette kan fx være som enzymer eller som proteiner der qua deres struktur kan danne komponenter der kan specifikke ting som fx at indgå i spindelvæv eller muskelfibre. Vi vil gerne aflure hvordan en specifik sekvens af aminosyrer danner en bestemt proteinstruktur. Det vil fortælle os en masse om hvilke protein de tusindvis af nye gener som identificeres hver eneste dag giver ophav til, men det vil også give os mulighed for at vende problemstillingen om og spørge: Givet denne struktur, hvilken sekvens af aminosyrer skal vi lave vha gensplejsning for at få dem til at folde op på den ønskede måde. Hvis vi kan forudsige dette vil vi på sigt kunne generere nye proteiner som kan ting som ikke kendes fra naturen. Det er bl.a. disse muligheder vi er i færd med at undersøge vha nogle simple biologiske modeller, bioinformatik og computertid : Evaluering ved Helle Blæsild og Karen Skriver samt diskussion med professor, Pierre Clément.. Frugt og drikkevarer i rum [] 3

4 Alkalisk fosfatase Introduktion I programpakkerne arbejdes med alkalisk fosfatase som et alternativt til de enzymer, der normalt benyttes til at bestemme steady-state enzymkinetiske parametre i gymnasiale undervisningsforløb. Dette er enzym er oplagt til gymnasieforsøg, fordi det er et meget stabilt enzym og har en interessant biologisk funktion. Ud over forsøg, der vil kunne udføres i gymnasiernes undervisningslokaler, præsenterer vi oplæg til programpakker, der vil kunne udføres som demonstrationsøvelser ved Biologisk Institut. Disse involverer bl.a. størrelsesbestemmelse vha. massespektrometri og stabilitetsbestemmelse vha. biofysiske analyser baseret på kalorimetri og fluorescensspektroskopi. Desuden kan mutantenzymer, dannet ved site-directed mutagenese, undersøges for aktivitet. Enzymet alkalisk fosfatase (AP) AP er et enzym, der katalyserer hydrolyse af fosforsyreestere, hvorved der dannes en alkohol- og en fosfatforbindelse (Fig. 1). Enzymet er kendt for at kunne fjerne fosfatgrupper i 5- og 3-positioner fra mange typer molekyler, blandt andet nukleotider. Det anvendes derfor i molekylærbiologi i forbindelse med arbejde med DNA, da disse ofte har fosfatgrupper i 5 positioner. Mængden af alkalisk fosfatase i blodet hos mennesker kan benyttes i diagnostisk sammenhæng. Fig. 1. Hydrolyse af en fosforsyreester vha. alkalisk fosfatase I de foreslåede forsøg oprenses AP fra en baterie, E. coli, i det AP findes i det periplasmatiske rum, uden for cellemembranen, i bakterier. Det gør oprensningen af enzymet nemmere, og enzymet har opnået stor grad af stabilitet for at overleve i dette miljø. E. coli AP er en dimer bestående af monomerer med en størrelse på ca. 49 kda. AP binder både zink- og magnesiumioner. ph optimum for AP fra E. coli er, som navnet antyder i det basiske område, og er 8,0 [] 4

5 Arbejdspakke 1 Produktion og oprensning af AP fra E. coli vha. osmotisk chok og varmebehandling* AP oprenses* fra det periplamatiske rum i E. coli stamme HO1064 (HO1064: F - aracam arad DE(lac)U169 trpam malam rpsl rela thi deod gsk-3 udp supf pstscab-phou ). Protokoller Vækst og oprensning 1) Pod 10 ml Luria Broth medium med en HO1064 koloni. Lad kulturen gror natten over ved 37 C. 2) Overfør 1 ml kultur til 1 l Luria Broth i en 5 l kolbe. Lad kulturen gro ved 37 C, 180 r.p.m., til sen stationær fase (ca. 28 timer). 3) Høst cellerne ved centrifugering 8000 r.p.m. i 10 min. Beregn celleudbyttet ved vejning. 4) Vask cellerne i 300 ml 10 mm Tris-HCl, 10 mm MgCl 2, ph 7,4. Centrifuger 8000 r.p.m. i 10 min og hæld supernatanten fra. Gentag 3 gange. 5) Resuspender cellerne i 60 mm Tris-HCl, ph 7,4 (1 gram pr. 10 ml) og tilsæt samme volumen 40 % sukrose. Tilsæt etylen-diamin-tetra-eddikesyre til 5 mm. 6) Overfør suspensionen til en større kolbe og ryst i 10 min. ved stue temperatur. 7) Centrifuger 15 min r.p.m. i 10 min, hæld supernatanten fra og placer bundfaldet på is. 8) Resuspender cellerne, som holdes på is, forsigtigt i 2 ml 40 % sukrose. 9) Tilsæt kold H 2 O (2 gange volumen af tilsat 60 mm Tris-HCl, ph 7,4). Herved frigives AP fra cellerne. 10) Overfør suspensionen til en ren kolbe og inkuber ved 4 C, 200 r.p.m. i 10 min. 11) Centrifuger 8000 r.p.m. i 15 min. og gem supernatanten. 12) Mål volumen af supernatanten og tilsæt Tris-HCl, ph 7,4 til 10 mm og MgCl 2 til 10 mm. Gem 1 ml til senere brug. 13) Udfæld proteinet ved langsomt (over 30 min.) at tilsætte ammoniumsulfat til 80 % mætning ved 4 C under omrøring. Lad opløsningen stå under omrøring i yderligere 30 min. 14) Centrifuger ammoniumsulfat bundfaldet ned ( r.p.m., 10 min) og resuspender i 50 ml TMZP (10 mm Tris-HCl, 1 mm MgCl 2, 10 μm ZnSO 4, 100 μm NaH 2 PO 4, - indstil ph til 7,4 med HCl). 15) Dialyser mod 1 l TMZP. 16) Fordel dialysatet i 1,5 ml rør og opvarm til 80 C på en varmeblok. 17) Centrifuger r.p.m. i 5 min. 18) Analyser oprensningsforløbet ved natriumdodecylsulfat (SDS)- polyakrylamidgelelektroforese (PAGE). *AP kan også fås i oprenset form ved henvendelse til en af ovennævnte kontaktpersoner Gelelektroforese Proteinoprensningsforløb og renhedsgraden af proteinpræparationer undersøges som regel ved gelelektroforese. Molekyler med en nettoladning vil vandre i et elektrisk felt i retning mod den elektrode, der har modsat retning af molekylets nettoladning. Den elektroforetiske mobilitet afhænger af molekylets nettoantal af ladede grupper og dets størrelse. Elektroforese til analyse af [] 5
. 3) Høst cellerne ved centrifugering 8000 r.p.m. i 10 min. Beregn celleudbyttet ved vejning.")
6 proteiner udføres som regel i et stabiliserende medium, en gel, der består af polyakrylamid. Polyakrylamidgeler fremstilles ved kopolymerisering af akrylamid og N,N -methylenbisakrylamid, der fungerer som krydsbinder. Natrium dodecylsulfat (SDS) polyakrylamidelektroforese (PAGE) kan benyttes til at estimere proteiners molekylvægt. Proteinet denatureres ved kogning med en opløsning af SDS, der er et anionisk detergent. Derefter separeres proteinerne i en polyakrylamidgel. Et protein denatureres, d.v.s. at den rumlige struktur ødelægges, når det koger i en opløsning af SDS. Proteinet vil samtidig få maskeret sine ladninger, fordi SDS bindes til det. Da SDS bindes til proteinet i et konstant forhold (1,4 gram SDS per gram protein) udjævnes ladningsforskelle, der skyldes sidekædernes ionisering. Resultatet bliver, at separationen vha. elektroforese kun afhænger af proteinets længde. Ved elektroforese i en polyakrylamidgel med 0,1 procent SDS vil der være en lineær sammenhæng mellem logaritmen til molekylvægten og den relative vandringslængde for protein-sds-komplekset. Molekylvægten kan bestemmes ved at behandle en række markørproteiner med kendt molekylvægt på samme måde og konstruere en standardkurve på grundlag af disse markører (Fig. 2). Fig. 2 Bestemmelse af molekylvægt ved SDS-PAGE. Man kan benytte forskellige støbestande og gelkar til SDS-PAGE. Ved Sektion for Biomolekylære Videnskaber benyttes et system fra BioRad ( yselector). [] 6

7 Arbejdspakke 2 Alkalisk phosphatase fra Escherichia coli enzymkinetik Phosphorylering og dephosphorylering af proteiner og stofskifte intermediater er udbredt i alle organismer og har i mange tilfælde vigtige regulatoriske funktioner (fx signaltransduktionsveje). Da dephosphorylering er kritiske reaktioner i cellesignalerings processer er phosphataser involveret i en lang række sygdomme. Her anvendes alkalisk phosphatase (AP) fra den gram negative bakterie Escherichia coli (E. coli) som katalyserer hydrolyse af phosphorsyreestre. ZN-ATOMER ER NØDVENDIGE FOR DEN ENZYMATISKE AKTIVITET. Dette enzym har en række fordele som model enzym. Det eksporteres gennem den indre membran ud i det periplasmatiske rum og kan derfor adskilles fra bakterierne ved at udsætte dem for et osmotisk shock. Enzymet er i modsætning til de fleste andre E. coli proteiner varmestabilt, hvilket gør oprensningen lettere og det kan derfor oprenses på de fleste gymnasier uden krav til avanceret apparatur. Enzymaktiviteten er let at bestemme og kan i den simpleste version bestemmes uden brug af et spektrofotometer. AP følger Michaelis-Menten kinetik og hæmmes kompetitivt af produktet uorganisk fosfat (HPO 4 2-, P i ) hvilket gør den velegnet til at illustrere basal enzymkinetik. Lidt om Spektroskopi (fra ref 1) En farvet opløsning indeholder kemiske forbindelser, som absorberer synligt lys ved bestemte bølgelængder. Dette udnyttes ved spektrofotometri, hvor der måles hvor meget lys ved en bestemt bølgelængde, der kan passere igennem en opløsning i forhold til en referenceopløsning. Størrelsen der måles, kaldes som bekendt absorbans. Den angives med et A og har ingen enhed. Absorbansen ved en bestemt bølgelængde afhænger af lysvejen (l i cm), koncentrationen af stoffet (her i mm) og en konstant ε λ som kaldes den molare absorptionskoefficient (ekstinktionskoefficienten, enhed fx M -1 cm -1 ). Sammenhængen kaldes Lambert-Beers lov: A = ε λ l [stof] Absorptionskurven har et karakteristisk forløb for det pågældende stof, og det er især interessant at vide, for hvilken bølgelængde den største absorption findes. Produktet i denne øvelse para-nitro-phenolation (pnp) har et absorptionsmaximum ved bølge-længden 405 nm, hvor den molare absorptions-koefficient ε λ er lig med 1850 M -1 cm -1 (talværdi stammer fra målinger foretaget på Panum Instituttet). Ved 410 nm er ε 410, ph8 (pnnp pnp) = 1620 M -1 cm -1 (ref 2). Lysvejen i forsøget er 1 cm. Med Lambert-Beers lov i hånden kan koncentrationen af reaktionsproduktet P beregnes, når [] 7

8 absorptionen måles, eller som vi er interesseret i dette forsøg, koncentrations-ændringen pr tid kan bestemmes ud fra ændringer i absorptionen pr tid : V = d[p]/dt = (1/ε λ l) da/dt Reaktionshastighed Michaelis-Menten enzymkinetik (ref 1) For simple enzymreaktioner har man iagttaget to karakteristiske egenskaber; i) når substratkoncentrationen er forholdsvis lille, så følger omdannelsen til produkt ofte, hvad der svarer til en første ordens reaktion med hensyn til substratkoncentrationen, og ii) når substratkoncentrationen er forholdsvis høj, så følger omdannelsen til produkt ofte, hvad der svarer til en nulte ordens reaktion med hensyn til substratkoncentrationen. I 1913 fremsatte Leonor Michaelis og Maud Menten en model for enzymreaktioner, som kunne forklare disse iagttagelser. Modellen fører frem til den såkaldte Michaelis-Menten ligning, i hvilken der indgår nogle størrelser, som kan karakterisere enzymreaktionen, nemlig Michaelis-Menten konstanten, K m, den maximale reaktionshastighed V max og hastighedskonstanten k cat, også kaldet enzymets turnover number. Michaelis-Menten ligningen benyttes stadig til at beskrive mange enzymers reaktionsforløb. I denne øvelse skal de tre konstanter K m, V max og k cat bestemmes for enzymet alkalisk phosphatase. I parentes bemærket kan nævnes at i midten af 1940 erne arbejdede Maud Menten også med dette enzym. Den enzymkatalyserede reaktion kan kort beskrives ved tre reaktioner. Substratet, S, bindes til enzymet, E, og der dannes et enzym-substrat kompleks, ES. Komplekset kan enten gå i stykker igen, hvorved substratet og enzymet gendannes, eller det kan omdannes til produktet P og enzymet. Til hver af de tre reaktioner er knyttet en hastighedskonstant. k 1 k cat E + S ES E + P k -1 Hvis det antages: i) substratkoncentrationen er meget større end enzymkoncentrationen, ii) omdannelsen af enzym-substrat kompleks til produktet og enzymet er hastighedsbestemmende og iii) koncentrationen af enzym-substrat komplekset ikke ændrer sig, kan Michaelis-Menten ligningen udledes: Fig 1 Ligningen viser sammenhængen substratkoncentrationen og reaktionshastigheden (se figur 3). For Michaelis-Menten konstanten, K m, den maximale reaktionshastighed V max og hastighedskonstanten k cat, gælder følgende ([E] o er enzymkoncentrationen): K m = (k -1 k cat )/k 1 og V max = k cat [E] 0 Hvis hastigheden V afbildes som funktion af substratkoncentrationen (fx begyndelses-koncentrationen), vil forløbet blive som det ses på figur 1. Grafen kan benyttes til at bestemme K m og V max. Der findes dog andre metoder, som er bedre kan benyttes til dette formål. [] 8

9 I dette forsøg findes typiske værdier for Michaelis-Menten konstanten, K m, omkring 0,15 µm, mens V max afhænger meget af enzymkoncentrationen, fra omkring 3 μm/min ved lave enzymkoncentrationer til omkring 10 μm/min ved høje enzymkoncentrationer. Det skal nævnes, at det langt fra er alle enzym-reaktioners substratafhængighed, som kan beskrives ved Michaelis-Menten modellen. Ved at bestemme initialhastigheden ved en række forskellige substratkoncentrationer kan en mætningskurve med aktivitet (V o ) som funktion af [S] tegnes. K m og V max kan bestemmes ved at fitte datapunkterne i et kurvefitteprogram til Michaelis-Menten ligningen. Man kan også tage det reciprokke af Michaelis-Menten ligningen hvorved fås: Fig 2 Når 1/V 0 afbildes som funktion af 1/[S] fås en ret linje med hældningen lig med K m /V max og afskæring med y-aksen lig med 1/V max (se figur 2). Hæmning af enzymaktiviteten Enzymers aktivitet kan påvirkes af forskellige hæmning. I dette tilfælde kan AP aktiviteten hæmmes af dets eget produkt P i, som er en kompetitiv hæmmer, dvs P i og substrat (i dette forsøg pnnp) konkurrerer om bindingen til det frie enzym. Dette påvirker K m mens V max ikke ændres. Michaelis-Menten ligningen for kompetitiv hæmning ser således ud:, og hvor Ved at måle initialhastigheder ved forskellige hæmmer koncentrationer kan K i (dissociationskonstanten) bestemmes ved kurve-fitning til ovenstående ligning. Spektrofotometer: Cecil CE series Målinger Reaktionerne foregår direkte i spektrofotometer kuvetter, og reaktionsmedierne blandes direkte i disse. Indstilling af spektrofotometeret Bølgelængden indstilles på 410 nm ved at trykke på Go to, 410, enter (E). Kinetik: [] 9
![Ved at bestemme initialhastigheden ved en række forskellige substratkoncentrationer kan en mætningskurve med aktivitet (V o ) som funktion af [S] tegnes.](/docs-images/44/2503138/images/page_9.jpg "K m og V max kan bestemmes ved at fitte datapunkterne i et kurvefitteprogram til Michaelis-Menten ligningen.")
10 Kinetik programmet beregner reaktionshastigheder ved at måle hastigheden hvormed absorptionen ændrer sig som funktion af tiden og kan således måle initial hældninger. Kinetik programmet aktiveres ved først at trykke på Quant Kinetics Menu Kinetics Indtastning af parametrene i Kinetic programmet Den ønskede værdi for de forskellige parametre indtastes og accepteres med E. Taster man forkert før man har trykket E kan der slettes med C. Da man ikke på forhånd kender den ønskede værdi for alle parametrene må man gætte sig lidt frem. Man kan når forsøget er afsluttet, gå ind i Kinetics reprocess og ændre på alle parametre undtagen end time som ikke må være begrænsende. Vejledende værdier for startparametre: 1) Enter start time [m:s]: sættes altid til ) Enter time1 [m:s]: sættes typisk til ) Enter time 2 [m:s]: sættes typisk til ca. 2 min efter time 1 (dvs sættes til 02:30) 4) Enter end time [m:s]: Sættes typisk til ) Enter factor sættes altid til (1/ε 410 = μm cm) 6) Enter min Abs: sættes til ) Enter max Abs: sættes til ) Enter scale [s/cm]: Dette er skalaen på x-aksen. Sættes til 40 Spektrofotometeret nulstilles på kuvettens indhold ved at trykke på Zero før assayet startes. Reagenser 1 M Tris-HCl ph8 4 M NaCl 5 mm ZnCl 2 H 2 O (hvid farvekode) 2x Buffermix (200 mm Tris-HCl ph8, 1 M NaCl, 200 µm ZnCl 2 ) (gul farvekode) 15 mm pnpp (opløst i 10 mm Tris-HCl ph8) (grøn farvekode) 1 mm pnpp (fortyndet i 10 mm Tris-HCl ph8) (blå farvekode) 1 mm P i ph8 (rød farvekode) [] 10

11 Enzym Alkalisk phosphatase (AP) er oprenset fra E. coli HO1064 (F - aracam arad DE(lac)U169 trpam malam rpsl rela thi deod gsk-3 udp supf pstscab-phou) (ref 3). Enzymet ligger i TMZP buffer (10 mm Tris-HCl, 1 mm MgCl 2, 10 μm ZnSO 4, 100 μm NaH 2 PO 4 ph7.4). Det udleverede enzym er fortyndet i 10 mm Tris-HCl ph8.0 til en koncentration på 2,4 µg/ml (0.05 nm) (koncentrationen er estimeret udfra en SDS polyacrylamidgel og absorbtion ved 280 nm). Det aktive enzym består af to identiske subunits (hver på 471 aminosyrer og Mr= 49434g/mol). Normalt måles enzymaktiviteter ved 25 C (eller 37 C), men i denne øvelse måles der ved stuetemperatur da de fleste gymnasier sandsynligvis ikke har termostaterede spektrofotometre. [] 11
.")
12 A. Bestemmelse af K m og V max µl µl µl µl Aktivitet Assay H 2 O 2x mix pnpp pnpp [pnpp] (15 mm) (1 mm) µm ΔA410/min µm/min ) Mix vand, buffermix og pnpp direkte i cuvetten og nulstil spektrofotometret. 2) Start assayet ved at tilsætte 10 µl AP (50 ng), mix ved omrøring og straks derefter starte målingen ved at trykke på RUN. 3) Efter et par minutter kan reaktionen stoppes og hældningen (initialhastigheden) kan bestemmes ved at t1 og t2 lægges på et passende sted hvor kurven er linear. Databehandling: Ved at afbilde enzym aktiviteten (fx µm produkt dannet per min eller blot ΔA410/min) som funktion af substratkoncentrationen fås en mætningskurve. K m og V max kan bestemmes direkte ved at punkterne fittes til Michealis-Menten ligningen i et kurve-fitting program eller, i mangel af software, bestemmes ud fra et et Lineweaver-Burk plot hvor 1/V afbildes som funktion af 1/[S]. [] 12
 Efter et par minutter kan reaktionen stoppes og hældningen (initialhastigheden) kan bestemmes ved at t1 og t2 lægges på et passende sted hvor kurven er linear.")
13 B. Bestemmelse af K m(app), V max(app) K I(Pi) Ved [P i ] = 50 µm µl µl µl µl µl Aktivitet Assay H 2 O 2x mix P i pnpp pnpp [pnpp] (1 mm) (15 mm) (1 mm) µm ΔA410/min µm/min ) Mix vand, buffermix, pnpp og P i direkte i cuvetten og nulstil spektrofotometret. 2) Start assayet ved at tilsætte 10 µl AP (50 ng), mix ved omrøring og straks derefter starte målingen ved at trykke på RUN. 3) Efter et par minutter kan reaktionen stoppes og hældningen (initialhastigheden) kan bestemmes ved at t1 og t2 lægges på et passende sted hvor kurven er linear. [] 13
 Start assayet ved at tilsætte 10 µl AP (50 ng), mix ved omrøring og straks derefter starte målingen ved at trykke på RUN.")
14 C. Bestemmelse af K m(app), V max(app) K I(Pi) Ved [Pi] = 100 µm µl µl µl µl µl Aktivitet Assay H 2 O 2x mix P i pnpp pnpp [pnpp] (1 mm) (15 mm) (1 mm) µm ΔA410/min µm/min ) Mix vand, buffermix, pnpp og P i direkte i cuvetten og nulstil spektrofotometret. 2) Start assayet ved at tilsætte 10 µl AP (50 ng), mix ved omrøring og straks derefter starte målingen ved at trykke på RUN. 3) Efter et par minutter kan reaktionen stoppes og hældningen (initialhastigheden) kan bestemmes ved at t1 og t2 lægges på et passende sted hvor kurven er linear. [] 14
 Start assayet ved at tilsætte 10 µl AP (50 ng), mix ved omrøring og straks derefter starte målingen ved at trykke på RUN.")
15 Øvrige forsøg (protokoller under udvikling): Det er let at udvide karakteriseringen med en lang række forsøg. Her er det nok at lave enkelt punkts asssays dvs assays ved høj substratkoncentration hvor aktiviteten ikke påvirkes af at en del af substratet forbruges. 1) AP kræver en Zn 2+ ion for at være aktiv. Her er AP opbevaret i en buffer med ZnCl 2, men det er let at fjerne Zn 2+ vha overskud af EDTA. Kinetikken af både inaktivering (med EDTA) og reaktivering (med overskud af ZnCl 2 ) kan let bestemmes. 2) Aktiviteten af AP afhænger af ionstyrken hvilket kan undersøges ved at ændre NaCl koncentration i en række assays. 3) ph optimum kan bestemmes vha en række buffere med forskelligt ph. Bemærk at ionstyrken af fx Tris-HCl buffer afhænger stærkt af ph. Dette kan udregnes og ionstyrken kan holdes konstant ved at ændre NaCl koncentrationen. 4) AP er temmelig varmestabil og bliver under oprensningen opvarmet til 80 C i 20 minutter. Varmestabiliteten af AP kan måles ved at inkubere enzymet ved fx 95 C. Ved at udtage prøver til forskellige tider og måle aktiviteten han en halveringstid bestemmes [] 15
 Aktiviteten af AP afhænger af ionstyrken hvilket kan undersøges ved at ændre NaCl koncentration i en række assays. 3) ph optimum kan bestemmes vha en række buffere med forskelligt ph.")
16 Arbejdspakke 3 Strukturanalyser af Alkalisk Fosfatase [] 16
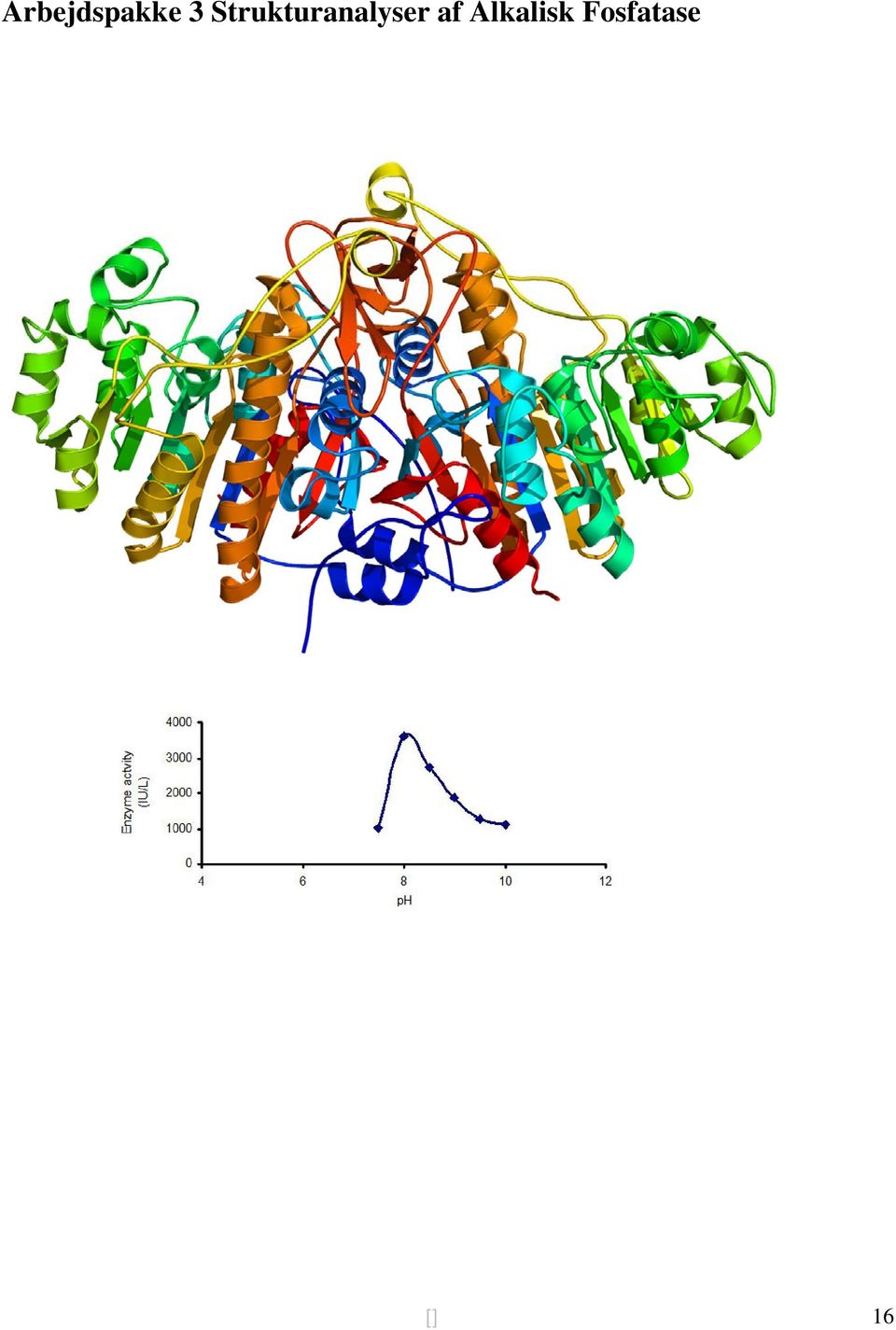
17 Introduktion til Struktur og Funktion af Alkalisk Fosfatase Enzymer er fantastiske mikromaskiner, hvis atomare strukturer udgør finmekanikken i selve processen. For at forstå hvorledes et enzym virker og for at kunne ændre denne funktion ved hjælp af proteindesign, er det vigitgt at kende dets struktur i detaljer. Der findes kun to metoder, hvormed man kan opnå denne information, nemlig krystallogfrafi og røngten diffraktion, samt kerne magnetisk resonans (NMR) spektroskopi. Begge disse metoder frembringer koordinater (x,v,z) for atomerne i enzymet (og andre biomolekyler) og disse kan visualiseres i dertil fremstillede programmer, de såkaldte molekylære viewers. For alkalisk fosfatase (AP) er der løst en stort sæt af strukturer og en søgning i strukturdatabasen giver mere en 60 hits. For alkalisk fosfatase fra E. coli nedbringes antallet lidt, men der er stadig mange at vælge imellem. Spørgsmået er, hvilken struktur man bedst er tjent med at vælge som analyseplatform. Det afhænger i høj grad af formålet med analysen og målet for ens design. Typisk findes der strukturer af forskellige varianter af enzymet (mutanter) og strukturer af komplekser med forskelllige ligander og inhibitorer. Vi har udvalgt et minimeret basissæt, som vil tjene som råmateriale for dagens in silico analyse. Vi skal gennem strukturanalyser af AP allerførst lære at bruge selve programmet og derigennem opnå forståelse for proteiners struktur. Derefter vil vi prøve at designe et sæt af enzymevarianter, som vi vil forvente enten vil have forskellige egenskaber i form af øget/sænket enzymaktivitet, ændret ph optimum eller ændret protein stabilitet. For at kunne lave nogle kvalificerede valg og ikke arbejde gennem trial and error analyser, anvender man ofte in silico værktøjer. Vi vil introducere to af disse værktøjer på kurset, PROPKA, der er et værktøj som udfra en strukturfil, kan beregne pka værdier for alle ioniserbare grupper i proteinet (input PDB fil). Det andet værktøj er ERIS som forudsiger ændringer i proteinet globale stabilitet på baggrund af en mutation. Dette værktøj arbejder også udfra en strukturfil. Begge programmer beskrives kort til sidst i denne oversigt. I øvelsen vil vi først give en kort introduktion til en strukturfil (PDB fil), til selve analyseprogrammerne (PyMOL, PROPKA, ERIS), og til hvorledes I kan løse dagens opgave. Der foreligger ikke et egentligt facit for selve designprocessen. Altså er udvælgelsen af, hvilken type variant af alkalisk fosfatase I ønsker at frembringe, helt jeres valg. Vi vil naturligvis være tilstede under øvelsen og være behjælpelig med programmerne. Rigtig god fornøjelse Kaare Teilum og Birthe Kragelund [] 17
 for atomerne i enzymet (og andre biomolekyler) og disse kan visualiseres i dertil fremstillede programmer, de såkaldte molekylære viewers.")
18 OPGAVEARK Følgende opgaver og analyser kan gennemføres på workshopdagen 1) Beskriv den overordnede struktur 2) Vis aminosyrerester i det aktive site (indenfor 5 Å) af PO4 i 1ED8 wt med PO ) Identificer ligander til PO 4 3, Mg 2+, Zn 2+ i 1ED8, og undersøg de samme rester i 1ED9 hvor der ikke er PO 4 3 bundet. Er der forskelle og ligheder? 4) Forklar hvordan Ser102 er aktiveret for nucleofilt angreb af en fosfoester. 5) Foreslå punktmutationer i det aktive site eller andre steder der vil a) ødelægge aktiviteten b) ændre ph optimum c) ændre proteinets stabilitet [] 18
 Forklar hvordan Ser102 er aktiveret for nucleofilt angreb af en fosfoester.")
19 Oversigt over tilgængelige strukturer af alkalisk fosfatase denne table indeholder oversigt over de tre strukturfiler, der skal udgøre basis for øvelsen PDB kode Molekyle Kommentarer 1ED8 Alkalisk fosfatase E. coli Inhiberet med uorganisk fosfat PO4 1ED9 Alkalisk fosfatase E. coli Uinhiberet Indeholder også magnesium, zink og sulfationer Indeholder også magnesium, zink og sulfationer 3DPC Alkalisk fosfatase E. coli mutantprotein I kompleks med fosforpeptid [] 19

20 Introduktion til PDB filer Hvad er PDB? The Protein Data Bank (PDB) er et arkiv (en database) over eksperimentelt bestemte tredimensionale strukturer af biologiske makromolekyler. Databasen anvendes globalt af forskere, undervisere og studerende. De data, der er indeholdt i databasen, er molekylernes atomare koordinater (rumlige placering (x,y,z) for hvert atom), bibliografiske citationer, den primære struktur (sekvensen), sekundær struktur information, krystallografiske struktur faktorer og eksperimentelle NMR data. PDB serverne The Worldwide Protein Data Bank (wwpdb) er en organisation, der systematiserer deponering, data processering og fordelingscentre for PDB data. Grundlæggerne er RCSB PDB (USA), MSD EBI (Europa) og PDBj (Japan). BMRB (USA) gruppen kom med I wwpdb i Missionen for wwpdb er til stadighed at bibeholde et eneste globalt arkiv over makromolekylære strukturelle data, gratis og frit tilgængeligt for hele den globale verden. Antallet af tilgængelige strukturer i PDB databasen stiger hele tiden. Den 22 december 2009 var antallet strukturer. En struktur er repræsenteret af en fil og alle strukturer har et firekarakters navn, feks. 1RW5. Nyttige links: PDB fil formatet En PDB fil er en helt almindelig tekstfil! Du kan læse den med hvilken teksteditor du har tilgængelig. De første linier i en pdb fil ser nogenlunde såden her ud: HEADER PROTEIN BINDING 19 NOV 03 1RJH TITLE STRUCTURE OF THE CALCIUM FREE FORM OF THE C TYPE LECTIN TITLE 2 LIKE DOMAIN OF TETRANECTIN og dette fortsætter de næste linier Der findes ligeledes en stort antal af programmer, der kan læse pdb filformatet, og vise en 3 dimensinal struktur på skærmen (se nedenfor). Record Format Enhver PDB fil kan ses som et antal af liner afsluttet med en end of line indikator. Hver linie består af 80 kolonner og er derfor også selvidentificerende. De føste seks kolonner af hver linie besåt af [] 20

21 et navn, ventrestillet og blank fyldt. Dette navn skal være et eksakt match med at af de navne der findes i databasen. PDB filen kan også ses som en samling af filer. Hver type fil indeholder en eller flere linier. Hver fil kan ydermere indeles i sektioner. Sektioner Tabellen nedenfor opsummerer de forskellige sektioner i en PDB fil og de forskellige felter (record types) de består af: Section Title: Summary descriptive remarks Remark: Bibliography, refinement Primary Structure: Peptide and/or nucleotide sequence and the relationship between the PDB sequence and that found in the sequence database(s) Heterogen: Non standard HET groups Secondary structure Connectivity: Chemical connectivities Miscellaneous features: Features within the macromolecule Crystallographic: Crystallographic cell Coordinate transformation: Coordinate transformation operators Coordinate: Atomic coordinate data Connectivity: Chemical connectivity Bookkeeping: Summary information, end of file marker Record Types HEADER, OBSLTE, TITLE, CAVEAT, COMPND, SOURCE, KEYWDS, EXPDTA, AUTHOR, REVDAT, SPRSDE, JRNL REMARKs 1, 2, 3 & annotations DBREF, SEQADV, SEQRES, MODRES HET, HETNAM, HETSYN, FORMUL HELIX, SHEET, TURN SSBOND, LINK, HYDBND, SLTBRG, CISPEP SITE CRYST1 ORIGXn, SCALEn, MTRIXn, TVECT MODEL, ATOM, SIGATM, ANISOU, SIGUIJ, TER, HETATM, ENDMDL CONECT MASTER, END Titelsektion Denne sektion indeholder de felter, der bruges til at beskrive eksperimentet og det biologiske makromolekyle for denne PDB ingang HEADER: Uniquely identifies a PDB entry through the idcode field. This record also provides a classification for the entry. Finally, it contains the date the coordinates were deposited at the PDB. o HEADER PROTEIN BINDING 19 NOV 03 1RJH TITLE: Contains a title for the experiment or analysis that is represented in the entry. It should identify an entry in the PDB in the same way that a title identifies a paper. COMPND: Describes the macromolecular contents of an entry. SOURCE: Biological and/or chemical source of each biological molecule in the entry. KEYWDS: Set of terms relevant to the entry. EXPDTA: Information about the experiment. AUTHOR: The names of the people responsible for the contents of the entry. REVDAT: History of the modifications made to an entry since its release. JRNL: Primary literature citation that describes the experiment which resulted in the deposited coordinate set. AUTH: List of authors associated with the cited article or contribution to a larger work TITL: Title of the reference. [] 21
[BESØGSSERVICE INSTITUT FOR MOLEKYLÆRBIOLOGI OG GENETIK, AU]
![[BESØGSSERVICE INSTITUT FOR MOLEKYLÆRBIOLOGI OG GENETIK, AU]](/thumbs/72/66305306.jpg "[BESØGSSERVICE INSTITUT FOR MOLEKYLÆRBIOLOGI OG GENETIK, AU]") Enzymkinetik INTRODUKTION Enzymer er biologiske katalysatorer i alle levende organismer som er essentielle for liv. Selektivt og effektivt katalyserer enzymerne kemiske reaktioner som ellers ikke ville
Enzymkinetik INTRODUKTION Enzymer er biologiske katalysatorer i alle levende organismer som er essentielle for liv. Selektivt og effektivt katalyserer enzymerne kemiske reaktioner som ellers ikke ville
[BESØGSSERVICE INSTITUT FOR MOLEKYLÆRBIOLOGI OG GENETIK, AU]
![[BESØGSSERVICE INSTITUT FOR MOLEKYLÆRBIOLOGI OG GENETIK, AU]](/thumbs/39/19540330.jpg "[BESØGSSERVICE INSTITUT FOR MOLEKYLÆRBIOLOGI OG GENETIK, AU]") Enzymkinetik INTRODUKTION Enzymer er biologiske katalysatorer i alle levende organismer som er essentielle for liv. Selektivt og effektivt katalyserer enzymerne kemiske reaktioner som ellers ikke ville
Enzymkinetik INTRODUKTION Enzymer er biologiske katalysatorer i alle levende organismer som er essentielle for liv. Selektivt og effektivt katalyserer enzymerne kemiske reaktioner som ellers ikke ville
Som substrat i forsøgene anvender vi para nitrophenylfosfat, der vha. enzymet omdannes til paranitrofenol
 Enzymkinetik Introduktion I disse forsøg skal I arbejde med enzymet alkalisk fosfatase. Fosfataser er meget almindelige i levende organismer og er enzymer med relativt bred substrat specificitet. De katalyserer
Enzymkinetik Introduktion I disse forsøg skal I arbejde med enzymet alkalisk fosfatase. Fosfataser er meget almindelige i levende organismer og er enzymer med relativt bred substrat specificitet. De katalyserer
Studienummer: MeDIS Exam 2015. Husk at opgive studienummer ikke navn og cpr.nr. på alle ark, der skal medtages i bedømmelsen
 MeDIS Exam 2015 Titel på kursus: Uddannelse: Semester: Videregående biokemi og medicinudvikling Bachelor i Medis 5. semester Eksamensdato: 26-01-2015 Tid: kl. 09.00-11.00 Bedømmelsesform 7-trin Vigtige
MeDIS Exam 2015 Titel på kursus: Uddannelse: Semester: Videregående biokemi og medicinudvikling Bachelor i Medis 5. semester Eksamensdato: 26-01-2015 Tid: kl. 09.00-11.00 Bedømmelsesform 7-trin Vigtige
mens mange enzymkatalyserede reaktioner er to-substrat reaktioner (2):
:") KemiF2 Enzymkinetik Trypsin er en serinproteinase, der katalyserer hydrolyse af peptid- og esterbindinger, hvor Arg eller Lys leverer carbonylgruppen. Ved øvelsen bestemmes de kinetiske parameterværdier,
KemiF2 Enzymkinetik Trypsin er en serinproteinase, der katalyserer hydrolyse af peptid- og esterbindinger, hvor Arg eller Lys leverer carbonylgruppen. Ved øvelsen bestemmes de kinetiske parameterværdier,
Forsøgene udføres ved betingelserne: ph 7.6, 0.1 M phosphatpuffer, ved stuetemperatur.
 KemiF2 nzymkinetik Trypsin er en serinproteinase, der katalyserer hydrolyse af peptid- og esterbindinger, hvor Arg eller Lys leverer carbonylgruppen. Ved øvelsen bestemmes de kinetiske parameterværdier,
KemiF2 nzymkinetik Trypsin er en serinproteinase, der katalyserer hydrolyse af peptid- og esterbindinger, hvor Arg eller Lys leverer carbonylgruppen. Ved øvelsen bestemmes de kinetiske parameterværdier,
Spm. 1.: Hvis den totale koncentration af monomer betegnes med CT hvad er så sammenhængen mellem CT, [D] og [M]?
![Spm. 1.: Hvis den totale koncentration af monomer betegnes med CT hvad er så sammenhængen mellem CT, [D] og [M]?](/thumbs/32/15386991.jpg "Spm. 1.: Hvis den totale koncentration af monomer betegnes med CT hvad er så sammenhængen mellem CT, [D] og [M]?") DNA-smeltetemperaturbestemmelse KemiF2-2008 DNA-smeltetemperaturbestemmelse Introduktion Oligonucleotider er ofte benyttet til at holde nanopartikler sammen med hinanden. Den ene enkeltstreng er kovalent
DNA-smeltetemperaturbestemmelse KemiF2-2008 DNA-smeltetemperaturbestemmelse Introduktion Oligonucleotider er ofte benyttet til at holde nanopartikler sammen med hinanden. Den ene enkeltstreng er kovalent
Side 1 af 14. Eksamen: Bioinformatik It og Sundhed 27 Jan 2011 kl 9-13
 Side 1 af 14 Eksamen: Bioinformatik It og Sundhed 27 Jan 2011 kl 9-13 Navn: Studie nummer: Dette eksamenssæt vil også kunne ses som en pdf fil nederst på kursus-hjemmesiden udfor den sidste dag d. 27 Jan
Side 1 af 14 Eksamen: Bioinformatik It og Sundhed 27 Jan 2011 kl 9-13 Navn: Studie nummer: Dette eksamenssæt vil også kunne ses som en pdf fil nederst på kursus-hjemmesiden udfor den sidste dag d. 27 Jan
Test Canvas: Eksamen i BMB502 Januar 2012
 BMB502, Enzymer og membraner, efterår 11. f Tests, Surveys and Pools Tests Test Canvas : Eksamen i BMB502 Januar 2012 Edit Mode is: Test Canvas: Eksamen i BMB502 Januar 2012 Create Reuse Upload s Settings
BMB502, Enzymer og membraner, efterår 11. f Tests, Surveys and Pools Tests Test Canvas : Eksamen i BMB502 Januar 2012 Edit Mode is: Test Canvas: Eksamen i BMB502 Januar 2012 Create Reuse Upload s Settings
KemiF2 Enzymkinetik. Ved en overfladisk beskrivelse af denne type enzymkatalyseret reaktion, angives reaktionen som (4):
:") KemiF2 nzymkinetik Trypsin er en serinproteinase, der katalyserer hydrolyse af peptid- og esterbindinger, hvor Arg eller Lys leverer carbonylgruppen. Ved øvelsen bestemmes de kinetiske parameterværdier,
KemiF2 nzymkinetik Trypsin er en serinproteinase, der katalyserer hydrolyse af peptid- og esterbindinger, hvor Arg eller Lys leverer carbonylgruppen. Ved øvelsen bestemmes de kinetiske parameterværdier,
Enzymkemi H. C. Ørsted Ungdomslaboratorium Kemisk Institut Københavns Universitet august 2001
 Enzymkemi H.. Ørsted Ungdomslaboratorium Kemisk Institut Københavns Universitet august 2001 2 Indholdsfortegnelse Enzymkinetik Indledning...2 Teori:...2 Mekanismen...2 Reaktionshastigheden:...5 Fremgangsmåde:...7
Enzymkemi H.. Ørsted Ungdomslaboratorium Kemisk Institut Københavns Universitet august 2001 2 Indholdsfortegnelse Enzymkinetik Indledning...2 Teori:...2 Mekanismen...2 Reaktionshastigheden:...5 Fremgangsmåde:...7
Titel: OPLØSELIGHEDEN AF KOBBER(II)SULFAT. Litteratur: Klasse: Dato: Ark 1 af. Helge Mygind, Kemi 2000 A-niveau 1, s. 290-292 8/9-2008/OV
SULFAT. Litteratur: Klasse: Dato: Ark 1 af. Helge Mygind, Kemi 2000 A-niveau 1, s. 290-292 8/9-2008/OV") Fag: KEMI Journal nr. Titel: OPLØSELIGHEDEN AF KOBBER(II)SULFAT Navn: Litteratur: Klasse: Dato: Ark 1 af Helge Mygind, Kemi 2000 A-niveau 1, s. 290-292 8/9-2008/OV Formålet er at bestemme opløseligheden
Fag: KEMI Journal nr. Titel: OPLØSELIGHEDEN AF KOBBER(II)SULFAT Navn: Litteratur: Klasse: Dato: Ark 1 af Helge Mygind, Kemi 2000 A-niveau 1, s. 290-292 8/9-2008/OV Formålet er at bestemme opløseligheden
W6 - Model- og Tegningskomposition
 W6 - Model- og Tegningskomposition Introduktion Dette er en kort introduktion til de nye værktøjer for tegningskomposition i 3D. Du vil komme til at arbejde med Clip Volume, Dynamic Views, Saved Views,
W6 - Model- og Tegningskomposition Introduktion Dette er en kort introduktion til de nye værktøjer for tegningskomposition i 3D. Du vil komme til at arbejde med Clip Volume, Dynamic Views, Saved Views,
Kemi F2- Laboratorieøvelse nr. 9 Ulla Christensen, Biofysisk Kemi ENZYMKINETIK
 1 Kemi F2- Laboratorieøvelse nr. 9 Ulla Christensen, Biofysisk Kemi ENZYMKINETIK Formål: Steady state hastighedsmålinger og kinetisk analyse til undersøgelse af enzymkatalyseret reaktion. Der gøres rede
1 Kemi F2- Laboratorieøvelse nr. 9 Ulla Christensen, Biofysisk Kemi ENZYMKINETIK Formål: Steady state hastighedsmålinger og kinetisk analyse til undersøgelse af enzymkatalyseret reaktion. Der gøres rede
DNA origami øvelse 2013. DNA origami øvelse
 DNA origami øvelse Introduktion I denne øvelse bruger vi DNA origami teknikken til at samle en tavle af DNA med dimensioner på 70 nm x 100 nm. Tavlen dannes af et langt enkeltstrenget DNA molekyle, der
DNA origami øvelse Introduktion I denne øvelse bruger vi DNA origami teknikken til at samle en tavle af DNA med dimensioner på 70 nm x 100 nm. Tavlen dannes af et langt enkeltstrenget DNA molekyle, der
BIOTEKNOLOGI HØJT NIVEAU
 STUDENTEREKSAMEN 2007 2007-BT-1 BITEKNLGI HØJT NIVEAU Torsdag den 31. maj 2007 kl. 9.00 14.00 Sættet består af 1 stor og 2 små opgaver samt 1 bilag i 2 eksemplarer. Det ene eksemplar af bilaget afleveres
STUDENTEREKSAMEN 2007 2007-BT-1 BITEKNLGI HØJT NIVEAU Torsdag den 31. maj 2007 kl. 9.00 14.00 Sættet består af 1 stor og 2 små opgaver samt 1 bilag i 2 eksemplarer. Det ene eksemplar af bilaget afleveres
PCR (Polymerase Chain Reaction): Opkopiering af DNA
: Opkopiering af DNA") PCR (Polymerase Chain Reaction): Opkopiering af DNA PCR til at opkopiere bestemte DNA-sekvenser i en prøve er nu en af genteknologiens absolut vigtigste værktøjer. Peter Rugbjerg, Biotech Academy PCR (Polymerase
PCR (Polymerase Chain Reaction): Opkopiering af DNA PCR til at opkopiere bestemte DNA-sekvenser i en prøve er nu en af genteknologiens absolut vigtigste værktøjer. Peter Rugbjerg, Biotech Academy PCR (Polymerase
1. Installere Logger Pro
 Programmet Logger Pro er et computerprogram, der kan bruges til at opsamle og behandle data i de naturvidenskabelige fag, herunder fysik. 1. Installere Logger Pro Første gang du installerer Logger Pro
Programmet Logger Pro er et computerprogram, der kan bruges til at opsamle og behandle data i de naturvidenskabelige fag, herunder fysik. 1. Installere Logger Pro Første gang du installerer Logger Pro
Fra spild til penge brug enzymer
 Fra spild til penge brug enzymer Køreplan 01005 Matematik 1 - FORÅR 2010 Denne projektplan er udarbejdet af Per Karlsson og Kim Knudsen, DTU Matematik, i samarbejde med Jørgen Risum, DTU Food. 1 Introduktion
Fra spild til penge brug enzymer Køreplan 01005 Matematik 1 - FORÅR 2010 Denne projektplan er udarbejdet af Per Karlsson og Kim Knudsen, DTU Matematik, i samarbejde med Jørgen Risum, DTU Food. 1 Introduktion
27611 Eksamen Sommer 2007
 - Side 1 af 10-27611 Eksamen Sommer 2007 Dette sæt indeholder 4 opgaver. En online version af opgavesættet vil være tilgængeligt fra kursets lektionsplan, under selve eksamen (25. Maj 2007 klokken 9:00
- Side 1 af 10-27611 Eksamen Sommer 2007 Dette sæt indeholder 4 opgaver. En online version af opgavesættet vil være tilgængeligt fra kursets lektionsplan, under selve eksamen (25. Maj 2007 klokken 9:00
Manual til udvidet abonnement
 Manual til udvidet abonnement April 2009 info@bookscan.dk (alle tal er fiktive) 1 LOG PÅ s. 3 FORSIDEN s. 4 TOP 500 s. 5 FORMATER s. 7 TIMELINE OG TRENDED TIMELINE s. 8 CHART WITH PROMPTS s. 14 SKEMASÆTNING
Manual til udvidet abonnement April 2009 info@bookscan.dk (alle tal er fiktive) 1 LOG PÅ s. 3 FORSIDEN s. 4 TOP 500 s. 5 FORMATER s. 7 TIMELINE OG TRENDED TIMELINE s. 8 CHART WITH PROMPTS s. 14 SKEMASÆTNING
Vejledning til opbygning af hjemmesider
 Side 1 af 9 Vejledning til opbygning af hjemmesider Hvis du er inde på din klubs hjemmeside, fx på forsiden, kan du nu gå i gang med at redigere. For at få redigeringsværktøjet frem, skal du klikke på
Side 1 af 9 Vejledning til opbygning af hjemmesider Hvis du er inde på din klubs hjemmeside, fx på forsiden, kan du nu gå i gang med at redigere. For at få redigeringsværktøjet frem, skal du klikke på
Side 1 of 11. Kursus navn: Kursus nr Introduktion til Bioinformatik
 Side 1 of 11 Danmarks Tekniske Universitet Skriftlig prøve, den 22/1-2015 Kursus navn: Kursus nr. 27633 Introduktion til Bioinformatik Tilladte hjælpemidler: Alle "Vægtning" Angivet ved de individuelle
Side 1 of 11 Danmarks Tekniske Universitet Skriftlig prøve, den 22/1-2015 Kursus navn: Kursus nr. 27633 Introduktion til Bioinformatik Tilladte hjælpemidler: Alle "Vægtning" Angivet ved de individuelle
Danmarks Tekniske Universitet
 Side 1 of 14 Danmarks Tekniske Universitet Skriftlig prøve, den 21/1-2013 Kursus navn: Kursus nr. 27633 Introduktion til Bioinformatik Tilladte hjælpemidler: Alle "Vægtning" Angivet ved de individuelle
Side 1 of 14 Danmarks Tekniske Universitet Skriftlig prøve, den 21/1-2013 Kursus navn: Kursus nr. 27633 Introduktion til Bioinformatik Tilladte hjælpemidler: Alle "Vægtning" Angivet ved de individuelle
Det Naturvidenskabelige Fakultet. Introduktion til Blackboard (Øvelser) Naturvidenskabeligt Projekt 2006 Prøv at forske
 Naturvidenskabeligt Projekt 2006 Prøv at forske") Det Naturvidenskabelige Fakultet Introduktion til Blackboard (Øvelser) Naturvidenskabeligt Projekt 2006 Prøv at forske Indholdsfortegnelse Introduktion til Blackboard Content System...3 Øvelse 01 individuel:
Det Naturvidenskabelige Fakultet Introduktion til Blackboard (Øvelser) Naturvidenskabeligt Projekt 2006 Prøv at forske Indholdsfortegnelse Introduktion til Blackboard Content System...3 Øvelse 01 individuel:
Verniers spektrofotometer SPRT-VIS USB 650
 Verniers spektrofotometer SPRT-VIS USB 650 Bølgelængdeinterval: 350 nm 1000 nm, nøjagtighed: < 1 nm. Brug Logger Pro s nyeste udgaver (3.6.0 eller 3.6.1). Hent evt. opdateringer fra Verniers hjemmeside
Verniers spektrofotometer SPRT-VIS USB 650 Bølgelængdeinterval: 350 nm 1000 nm, nøjagtighed: < 1 nm. Brug Logger Pro s nyeste udgaver (3.6.0 eller 3.6.1). Hent evt. opdateringer fra Verniers hjemmeside
Protein databases Rasmus Wernersson. (Slides af Henrik Nielsen & Morten Nielsen).
.") Protein databases Rasmus Wernersson (Slides af Henrik Nielsen & Morten Nielsen). Background- Nucleotide databases GenBank, http://www.ncbi.nlm.nih.gov/genbank/ National Center for Biotechnology Information
Protein databases Rasmus Wernersson (Slides af Henrik Nielsen & Morten Nielsen). Background- Nucleotide databases GenBank, http://www.ncbi.nlm.nih.gov/genbank/ National Center for Biotechnology Information
BIOZYMER ØVELSE 1 ENZYMKINETIK
 BIZYME ØVELE 1 EZYMKIETIK FAGLIG BAGGUD Det forudsættes, at teorien bag enzymer og enzymkinetik er bekendt. ertil kan bl.a. henvises til kapitlet om antibiotika og resistens i Bioteknologiske orisonter,
BIZYME ØVELE 1 EZYMKIETIK FAGLIG BAGGUD Det forudsættes, at teorien bag enzymer og enzymkinetik er bekendt. ertil kan bl.a. henvises til kapitlet om antibiotika og resistens i Bioteknologiske orisonter,
Kvantitativ bestemmelse af reducerende sukker (glukose)
") Kvantitativ bestemmelse af reducerende sukker (glukose) Baggrund: Det viser sig at en del af de sukkerarter vi indtager med vores mad er hvad man i fagsproget kalder reducerende sukkerarter. Disse vil
Kvantitativ bestemmelse af reducerende sukker (glukose) Baggrund: Det viser sig at en del af de sukkerarter vi indtager med vores mad er hvad man i fagsproget kalder reducerende sukkerarter. Disse vil
KUNDE-WEBPORTAL KOM GODT I GANG
 Specialtrykkeriet Arco KUNDE-WEBPORTAL KOM GODT I GANG Vi håber du får fornøjelse af vores nye værktøj. WebApproval skal gøre det nemmere og mere sikkert for dig at godkende din tryksager. Har du spørgsmål,
Specialtrykkeriet Arco KUNDE-WEBPORTAL KOM GODT I GANG Vi håber du får fornøjelse af vores nye værktøj. WebApproval skal gøre det nemmere og mere sikkert for dig at godkende din tryksager. Har du spørgsmål,
Energi, Enzymer & enzymkinetik.metabolisme
 (gruppeopgaver i databar 152 (og 052)) Energi, Enzymer & enzymkinetik.metabolisme Tirsdag den 17. september kl 13-14.15 (ca) Auditorium 53, bygning 210 Susanne Jacobsen sja@bio.dtu.dk Enzyme and Protein
(gruppeopgaver i databar 152 (og 052)) Energi, Enzymer & enzymkinetik.metabolisme Tirsdag den 17. september kl 13-14.15 (ca) Auditorium 53, bygning 210 Susanne Jacobsen sja@bio.dtu.dk Enzyme and Protein
Bestemmelse af koffein i cola
 Bestemmelse af koffein i cola 1,3,7-trimethylxanthine Koffein i læskedrikke Læs følgende links, hvor der blandt andet står nogle informationer om koffein og regler for hvor meget koffein, der må være i
Bestemmelse af koffein i cola 1,3,7-trimethylxanthine Koffein i læskedrikke Læs følgende links, hvor der blandt andet står nogle informationer om koffein og regler for hvor meget koffein, der må være i
Amalie Avnborg 2.y SRO 18/3-12
 Side 1 af 23 SRO for 2. y 2012 Opgaveformulering: Med udgangspunkt i enzymet catecholase, som du har lavet to forsøg med, skal du lave enzymkinetiske undersøgelser. Du skal redegøre for centrale begreber
Side 1 af 23 SRO for 2. y 2012 Opgaveformulering: Med udgangspunkt i enzymet catecholase, som du har lavet to forsøg med, skal du lave enzymkinetiske undersøgelser. Du skal redegøre for centrale begreber
Dannelse af PDF-dokumenter
 Dannelse af PDF-dokumenter Indhold Generere PDF-dokumenter... 2 Håndtering af PDF-dokumentet... 8 Hvordan indsætter man sidetal i PDF-dokumentet?... 8 Hvordan laver man bookmarks i PDF-dokumentet?... 8
Dannelse af PDF-dokumenter Indhold Generere PDF-dokumenter... 2 Håndtering af PDF-dokumentet... 8 Hvordan indsætter man sidetal i PDF-dokumentet?... 8 Hvordan laver man bookmarks i PDF-dokumentet?... 8
Litteratursøgning i PubMed
 Litteratursøgning i PubMed 1 En introduktion til PubMed PubMed er National Library of Medicine s webadgang til Medline, produceret i samarbejde med tidsskriftudgiverne. Tidsskriftudgiverne leverer data
Litteratursøgning i PubMed 1 En introduktion til PubMed PubMed er National Library of Medicine s webadgang til Medline, produceret i samarbejde med tidsskriftudgiverne. Tidsskriftudgiverne leverer data
Dannelse af PDF dokumenter
 Dannelse af PDF dokumenter Indhold Dannelse af PDF-dokumenter i Phd Planner... 2 Valg af vedhæftninger i PDF dokumentet... 2 Valg af skabelon for PDF dokumentet... 3 Når PDF filen er dannet... 5 Gem PDF
Dannelse af PDF dokumenter Indhold Dannelse af PDF-dokumenter i Phd Planner... 2 Valg af vedhæftninger i PDF dokumentet... 2 Valg af skabelon for PDF dokumentet... 3 Når PDF filen er dannet... 5 Gem PDF
FORCE Inspect Online Manual v. 1.02. FORCE Inspect Online Manual. 1 af 18
 FORCE Inspect Online Manual 1 af 18 Indholdsfortegnelse Indholdsfortegnelse... 2 FORCE Inspect Online Manual... 3 Generelt... 3 Login... 3 Main... 4 Intro sektion... 4 Links sektion... 4 News sektion...
FORCE Inspect Online Manual 1 af 18 Indholdsfortegnelse Indholdsfortegnelse... 2 FORCE Inspect Online Manual... 3 Generelt... 3 Login... 3 Main... 4 Intro sektion... 4 Links sektion... 4 News sektion...
Side 1 af 13. Eksamen: Bioinformatik It og Sundhed 27 Jan 2011 kl 9-13
 Side1af13 Eksamen: Bioinformatik It og Sundhed 27 Jan 2011 kl 9-13 Navn: Studie nummer: Dette eksamenssæt vil også kunne ses som en pdf fil nederst på kursus-hjemmesiden udfor den sidste dag d. 27 Jan
Side1af13 Eksamen: Bioinformatik It og Sundhed 27 Jan 2011 kl 9-13 Navn: Studie nummer: Dette eksamenssæt vil også kunne ses som en pdf fil nederst på kursus-hjemmesiden udfor den sidste dag d. 27 Jan
Sådan opretter du en elektronisk aflevering
 Sådan arbejder du med opgaver i Gradebook/karakterbog Denne vejledning indeholder en detaljeret beskrivelse af hvordan du bruger gradebook/karakterbogen når du vil arbejde med opgaver og give karakterer
Sådan arbejder du med opgaver i Gradebook/karakterbog Denne vejledning indeholder en detaljeret beskrivelse af hvordan du bruger gradebook/karakterbogen når du vil arbejde med opgaver og give karakterer
Excel tutorial om lineær regression
 Excel tutorial om lineær regression I denne tutorial skal du lære at foretage lineær regression i Microsoft Excel 2007. Det forudsættes, at læseren har været igennem det indledende om lineære funktioner.
Excel tutorial om lineær regression I denne tutorial skal du lære at foretage lineær regression i Microsoft Excel 2007. Det forudsættes, at læseren har været igennem det indledende om lineære funktioner.
Rationel VinduesDesigner TM Brugervejledning
 Rationel VinduesDesigner TM Brugervejledning indhold: introduktion Side 2 Funktionsliste Side 3 Få adgang til systemet Side 4 opload dine billeder Side 5 Sådan bruges systemet Side 6 Gem dine eksempler
Rationel VinduesDesigner TM Brugervejledning indhold: introduktion Side 2 Funktionsliste Side 3 Få adgang til systemet Side 4 opload dine billeder Side 5 Sådan bruges systemet Side 6 Gem dine eksempler
DNA smeltepunktsbestemmelse
 DNA smeltepunktsbestemmelse Troels Linnet Christine Hartmann Mads Topp 29. november 2006 Resumé DNA smeltepunktet bestemmes teoretisk og praktisk til sammenligning. Ved opvarmning forventes et højere smeltepunkt
DNA smeltepunktsbestemmelse Troels Linnet Christine Hartmann Mads Topp 29. november 2006 Resumé DNA smeltepunktet bestemmes teoretisk og praktisk til sammenligning. Ved opvarmning forventes et højere smeltepunkt
Indholdsfortegnelse. Indholdsfortegnelse.. side 2. Adgang til webgraf 3. Opslag adresse... 4. Styring af layout.. 5. Zoom funktioner..
 Indholdsfortegnelse Indholdsfortegnelse.. side 2 Adgang til webgraf 3 Opslag adresse... 4 Styring af layout.. 5 Zoom funktioner.. 6 Panorere på skærmen. 7 Information om grafikken.... 8-10 Print et udsnit.....
Indholdsfortegnelse Indholdsfortegnelse.. side 2 Adgang til webgraf 3 Opslag adresse... 4 Styring af layout.. 5 Zoom funktioner.. 6 Panorere på skærmen. 7 Information om grafikken.... 8-10 Print et udsnit.....
1: Kemisk kinetik 1. Du skal gøre rede for kemiske reaktioners hastighed, herunder begrebet reaktionsorden.
 1: Kemisk kinetik 1. Du skal gøre rede for kemiske reaktioners hastighed, herunder begrebet reaktionsorden. Du skal gøre rede for eksperimentet: Krystalviolet. Du skal inddrage nogle af stikordene: Reaktionshastighed;
1: Kemisk kinetik 1. Du skal gøre rede for kemiske reaktioners hastighed, herunder begrebet reaktionsorden. Du skal gøre rede for eksperimentet: Krystalviolet. Du skal inddrage nogle af stikordene: Reaktionshastighed;
Lav din egen forside i webtrees
 Lav din egen forside i webtrees Du behøver ikke at kunne kode eller gøre noget advanceret for at designe din helt egen forside i webtrees. Alt du skal gøre er bare at gøre brug af den indbygget editor.
Lav din egen forside i webtrees Du behøver ikke at kunne kode eller gøre noget advanceret for at designe din helt egen forside i webtrees. Alt du skal gøre er bare at gøre brug af den indbygget editor.
Analyse af proteiner Øvelsesvejledning
 Center for Undervisningsmidler, afdeling København Analyse af proteiner Øvelsesvejledning Formål At separere og analysere proteiner i almindelige fødevarer ved brug af gelelektroforese. Teori Alle dele
Center for Undervisningsmidler, afdeling København Analyse af proteiner Øvelsesvejledning Formål At separere og analysere proteiner i almindelige fødevarer ved brug af gelelektroforese. Teori Alle dele
RefWorks en vejledning fra UCL Biblioteket. Indholdsfortegnelse
 Indholdsfortegnelse Hvad er RefWorks?... 2 Opret dig som bruger... 2 Inden du går i gang... 3 Klargøring af computer til download af Write-N-Cite v. 4.2... 3 Installation af Write-N-Cite... 4 Installation
Indholdsfortegnelse Hvad er RefWorks?... 2 Opret dig som bruger... 2 Inden du går i gang... 3 Klargøring af computer til download af Write-N-Cite v. 4.2... 3 Installation af Write-N-Cite... 4 Installation
Side%1%af%14% Eksamen: Bioinformatik It og Sundhed 27 Jan 2011 kl 9-13
 Side1af14 Eksamen: Bioinformatik It og Sundhed 27 Jan 2011 kl 9-13 Navn: Studie nummer: Dette eksamenssæt vil også kunne ses som en pdf fil nederst på kursus-hjemmesiden udfor den sidste dag d. 27 Jan
Side1af14 Eksamen: Bioinformatik It og Sundhed 27 Jan 2011 kl 9-13 Navn: Studie nummer: Dette eksamenssæt vil også kunne ses som en pdf fil nederst på kursus-hjemmesiden udfor den sidste dag d. 27 Jan
Vejledning KPK Online Prøverum
 Vejledning KPK Online Prøverum INDHOLD Introduktion side 2 Funktionsliste side 2 Få adgang til systemet side 3 Opload dine billeder side 4 Sådan bruges systemet side 5 Gem dine eksempler side 7 Side 1/7
Vejledning KPK Online Prøverum INDHOLD Introduktion side 2 Funktionsliste side 2 Få adgang til systemet side 3 Opload dine billeder side 4 Sådan bruges systemet side 5 Gem dine eksempler side 7 Side 1/7
Bioteknologi A. Studentereksamen. Af opgaverne 1 og 2 skal begge opgaver besvares. Af opgaverne 3 og 4 skal en og kun en af opgaverne besvares.
 Bioteknologi A Studentereksamen Af opgaverne 1 og 2 skal begge opgaver besvares. Af opgaverne 3 og 4 skal en og kun en af opgaverne besvares. frs111-btk/a-31052011 Tirsdag den 31. maj 2011 kl. 9.00-14.00
Bioteknologi A Studentereksamen Af opgaverne 1 og 2 skal begge opgaver besvares. Af opgaverne 3 og 4 skal en og kun en af opgaverne besvares. frs111-btk/a-31052011 Tirsdag den 31. maj 2011 kl. 9.00-14.00
Røntgenspektrum fra anode
 Røntgenspektrum fra anode Elisabeth Ulrikkeholm June 24, 2016 1 Formål I denne øvelse skal I karakterisere et røntgenpektrum fra en wolframanode eller en molybdænanode, og herunder bestemme energien af
Røntgenspektrum fra anode Elisabeth Ulrikkeholm June 24, 2016 1 Formål I denne øvelse skal I karakterisere et røntgenpektrum fra en wolframanode eller en molybdænanode, og herunder bestemme energien af
Protein syntese. return
 Protein syntese. I artiklen redegøres for principperne i, hvordan octapeptidet SCHTFGDI kan syntetiseres. Som yderligere illustration heraf kan peptidet opbygges og visualiseres i Chem3D-Pro. Herved kan
Protein syntese. I artiklen redegøres for principperne i, hvordan octapeptidet SCHTFGDI kan syntetiseres. Som yderligere illustration heraf kan peptidet opbygges og visualiseres i Chem3D-Pro. Herved kan
Når du holder møder i Connect
 Når du holder møder i Connect Det er vigtigt at den/de der er host og presenter på mødet sidder ved en forholdsvis kraftig computer, og har en god bredbåndsforbindelse. Hvis man skal vise præsentationer,
Når du holder møder i Connect Det er vigtigt at den/de der er host og presenter på mødet sidder ved en forholdsvis kraftig computer, og har en god bredbåndsforbindelse. Hvis man skal vise præsentationer,
Unitel EDI MT940 June 2010. Based on: SWIFT Standards - Category 9 MT940 Customer Statement Message (January 2004)
") Unitel EDI MT940 June 2010 Based on: SWIFT Standards - Category 9 MT940 Customer Statement Message (January 2004) Contents 1. Introduction...3 2. General...3 3. Description of the MT940 message...3 3.1.
Unitel EDI MT940 June 2010 Based on: SWIFT Standards - Category 9 MT940 Customer Statement Message (January 2004) Contents 1. Introduction...3 2. General...3 3. Description of the MT940 message...3 3.1.
Indholdsfortegnelse: SUPPORT Har du brug for hjælp til anvendelse af Qwickly Attendance eller Qwickly+, er du velkommen til at kontakte
 Qwickly er tilgængelig via Blackboard og består af to dele: Qwickly Attendance og Qwickly+. Qwickly Attendance håndterer fremmøderegistrering på de enkelte kurser, hvor Qwickly+ via forsiden af Blackboard
Qwickly er tilgængelig via Blackboard og består af to dele: Qwickly Attendance og Qwickly+. Qwickly Attendance håndterer fremmøderegistrering på de enkelte kurser, hvor Qwickly+ via forsiden af Blackboard
DNA origami øvelse DNA origami øvelse
 DNA origami øvelse Introduktion I denne øvelse bruger vi DNA origami teknikken til at samle en tavle af DNA med dimensioner på 70 nm x 100 nm. Tavlen dannes af et langt enkeltstrenget DNA molekyle, der
DNA origami øvelse Introduktion I denne øvelse bruger vi DNA origami teknikken til at samle en tavle af DNA med dimensioner på 70 nm x 100 nm. Tavlen dannes af et langt enkeltstrenget DNA molekyle, der
ADMINISTRATIONS MANUAL
 ADMINISTRATIONS MANUAL onmap.dk Administrations Manual Dansk Version 0.1 Side 1 Denne manual beskrive hvordan en race administrator kan opsætte og bruge onmap.dk race protalen til at lave en specialiseret
ADMINISTRATIONS MANUAL onmap.dk Administrations Manual Dansk Version 0.1 Side 1 Denne manual beskrive hvordan en race administrator kan opsætte og bruge onmap.dk race protalen til at lave en specialiseret
DNA origami øvelse. Introduktion. DNA origami øvelse 3 timer 2018
 DNA origami øvelse Introduktion I denne øvelse bruger vi DNA origami teknikken til at samle en tavle af DNA med dimensioner på 70 nm x 100 nm. Tavlen dannes af et langt enkeltstrenget DNA molekyle, der
DNA origami øvelse Introduktion I denne øvelse bruger vi DNA origami teknikken til at samle en tavle af DNA med dimensioner på 70 nm x 100 nm. Tavlen dannes af et langt enkeltstrenget DNA molekyle, der
MANUAL. Siteloom CMS
 MANUAL Siteloom CMS www.hjerteforeningen.dk/cms Brugernavn: Password: 3. september, 2012 BASIS FUNKTIONER 1. Kalender... 4 1.a. Opret... 5 1.b. Rediger eller slet... 8 2. Sider... 10 2.a Opret side...
MANUAL Siteloom CMS www.hjerteforeningen.dk/cms Brugernavn: Password: 3. september, 2012 BASIS FUNKTIONER 1. Kalender... 4 1.a. Opret... 5 1.b. Rediger eller slet... 8 2. Sider... 10 2.a Opret side...
Worldtrack Tracking Platform BRUGERVEJLEDNING Version 2.01
 2018 Worldtrack Tracking Platform BRUGERVEJLEDNING Version 2.01 WORLDTRACK Ejby industrivej 2, 2600 Glostrup Indhold Introduktion... 2 Login... 2 Menu... 2 Overvågning... 3 Bevægelses status... 4 GPS data
2018 Worldtrack Tracking Platform BRUGERVEJLEDNING Version 2.01 WORLDTRACK Ejby industrivej 2, 2600 Glostrup Indhold Introduktion... 2 Login... 2 Menu... 2 Overvågning... 3 Bevægelses status... 4 GPS data
Brug af Zeiss DuraMax
 Brug af Zeiss DuraMax Indhold Opstart af Duramax... 3 Brug af eksisterende Measurement Plan (Program)... 4 Opmåling af emne... 4 Udprintning... 6 Import af CAD model... 7 Klargør solid til måling... 8
Brug af Zeiss DuraMax Indhold Opstart af Duramax... 3 Brug af eksisterende Measurement Plan (Program)... 4 Opmåling af emne... 4 Udprintning... 6 Import af CAD model... 7 Klargør solid til måling... 8
27611 Eksamen Sommer 2008
 27611 Eksamen Sommer 2008 Dette sæt indeholder 10 opgaver. En online version af opgavesættet vil være tilgængeligt fra kursets lektionsplan under selve eksamen ( juni 2008 klokken 15:00-19:00). DNA/Protein
27611 Eksamen Sommer 2008 Dette sæt indeholder 10 opgaver. En online version af opgavesættet vil være tilgængeligt fra kursets lektionsplan under selve eksamen ( juni 2008 klokken 15:00-19:00). DNA/Protein
Simulering af en Mux2
 Simulering af en Mux2 Indhold Start QuartusII op start et nyt projekt.... 2 Fitter opsætning... 6 Opstart af nyt Block diagram... 8 ModelSim... 14 Hvis man vil ændre data grafisk kan det også lade sig
Simulering af en Mux2 Indhold Start QuartusII op start et nyt projekt.... 2 Fitter opsætning... 6 Opstart af nyt Block diagram... 8 ModelSim... 14 Hvis man vil ændre data grafisk kan det også lade sig
Hjælp, mine deltagere aflytter og øver sig til YouTube men i forkert toneart.
 Side 1 Gratis program til at transponere lydfil og gemme den, link og vejledning Ole Skou 2009 Hjælp, mine deltagere aflytter og øver sig til YouTube men i forkert toneart Gratis program til at transponere
Side 1 Gratis program til at transponere lydfil og gemme den, link og vejledning Ole Skou 2009 Hjælp, mine deltagere aflytter og øver sig til YouTube men i forkert toneart Gratis program til at transponere
Mendeley kan hjælpe dig med at organisere din forskning og samarbejde med andre online.
 Mendeley på Mac Mendeley er både en reference manager og et akademisk socialt netværk. Mendeley kan hjælpe dig med at organisere din forskning og samarbejde med andre online. Mendeley kan generere litteraturlister
Mendeley på Mac Mendeley er både en reference manager og et akademisk socialt netværk. Mendeley kan hjælpe dig med at organisere din forskning og samarbejde med andre online. Mendeley kan generere litteraturlister
Oprettelse af Titelblok i Capture og Capture CIS
 e-service Titelblok i OrCAD Capture og Capture CIS Side 1 af 11 Oprettelse af Titelblok i Capture og Capture CIS Note skrevet af : Nordcad Systems Technical Support Revision : April 2003, Release 14.2/9.2.3,
e-service Titelblok i OrCAD Capture og Capture CIS Side 1 af 11 Oprettelse af Titelblok i Capture og Capture CIS Note skrevet af : Nordcad Systems Technical Support Revision : April 2003, Release 14.2/9.2.3,
Picto Selector. Lav dine egne flotte symbolark på den nemme måde. Version: Oktober 2012
 Picto Selector Lav dine egne flotte symbolark på den nemme måde Version: Oktober 2012 Indholdsfortegnelse Hvad er Picto Selector?...4 USB?...4 Hent programmet...4 Installer programmet på din computer...5
Picto Selector Lav dine egne flotte symbolark på den nemme måde Version: Oktober 2012 Indholdsfortegnelse Hvad er Picto Selector?...4 USB?...4 Hent programmet...4 Installer programmet på din computer...5
SPECIALTRYKKERIETS KUNDE WEBPORTAL KOM GODT I GANG
 SPECIALTRYKKERIETS KUNDE WEBPORTAL KOM GODT I GANG Jeg håber du får fornøjelse af vores nye værktøj. WebApproval skal gøre det nemmere og mere sikkert for dig at godkende din tryksager. Har du spørgsmål,
SPECIALTRYKKERIETS KUNDE WEBPORTAL KOM GODT I GANG Jeg håber du får fornøjelse af vores nye værktøj. WebApproval skal gøre det nemmere og mere sikkert for dig at godkende din tryksager. Har du spørgsmål,
Baggrundsmateriale til Minigame 7 side 1 A + B C + D
 Baggrundsmateriale til Minigame 7 side 1 Indhold Kernestof... 1 Supplerende stof... 1 1. Differentialligninger (Baggrundsmateriale til Minigame 3)... 1 2. Reaktionsorden (Nulte-, første- og andenordensreaktioner)...
Baggrundsmateriale til Minigame 7 side 1 Indhold Kernestof... 1 Supplerende stof... 1 1. Differentialligninger (Baggrundsmateriale til Minigame 3)... 1 2. Reaktionsorden (Nulte-, første- og andenordensreaktioner)...
Laboratorieprotokol for manuel isolering af DNA fra 0,5 ml prøve
 Laboratorieprotokol for manuel isolering af DNA fra 0,5 ml prøve Til isolering af genomisk DNA fra indsamlingssætserierne Oragene - og ORAcollect. Du kan finde flere sprog og protokoller på vores webside
Laboratorieprotokol for manuel isolering af DNA fra 0,5 ml prøve Til isolering af genomisk DNA fra indsamlingssætserierne Oragene - og ORAcollect. Du kan finde flere sprog og protokoller på vores webside
Quick guide til e-learn.sdu.dk (Blackboard) for studerende
 for studerende") Quick guide til e-learn.sdu.dk (Blackboard) for studerende Der opnås adgang til e-learn.sdu.dk (Blackboard) via single sign-on, sso.sdu.dk/, eller adressen: e-learn.sdu.dk/. Når du har logget på, står
Quick guide til e-learn.sdu.dk (Blackboard) for studerende Der opnås adgang til e-learn.sdu.dk (Blackboard) via single sign-on, sso.sdu.dk/, eller adressen: e-learn.sdu.dk/. Når du har logget på, står
En forsker har lavet et cdna insert vha PCR og har anvendt det følgende primer sæt, som producerer hele den åbne læseramme af cdna et:
 F2011-Opgave 1. En forsker har lavet et cdna insert vha PCR og har anvendt det følgende primer sæt, som producerer hele den åbne læseramme af cdna et: Forward primer: 5 CC ATG GGT ATG AAG CTT TGC AGC CTT
F2011-Opgave 1. En forsker har lavet et cdna insert vha PCR og har anvendt det følgende primer sæt, som producerer hele den åbne læseramme af cdna et: Forward primer: 5 CC ATG GGT ATG AAG CTT TGC AGC CTT
Biotechnology Explorer. Protein Fingerprinting
 Biotechnology Explorer Protein Fingerprinting Instruktionsmanual Katalognummer 166-0100EDU explorer.bio-rad.com Delene i dette kit er sendt i seperate æsker. Opbevar proteinstandarderne i fryseren, ved
Biotechnology Explorer Protein Fingerprinting Instruktionsmanual Katalognummer 166-0100EDU explorer.bio-rad.com Delene i dette kit er sendt i seperate æsker. Opbevar proteinstandarderne i fryseren, ved
Mendeley kan hjælpe dig med at organisere din forskning og samarbejde med andre online.
 Mendeley på Mac Mendeley er både en reference manager og et akademisk socialt netværk. Mendeley kan hjælpe dig med at organisere din forskning og samarbejde med andre online. Mendeley kan generere litteraturlister
Mendeley på Mac Mendeley er både en reference manager og et akademisk socialt netværk. Mendeley kan hjælpe dig med at organisere din forskning og samarbejde med andre online. Mendeley kan generere litteraturlister
Abstract:... 1. Indledning til opgaven... 3. Introduktion til emnet... 4. Katalase generelt:... 6. Enzymers strukturelle opbygning...
 Abstract: This study has been made to describe enzymes, especially their kinetics and structures, and is based on the enzyme catalase. The kinetics has been explained by the Michaelis-Menten-equation and
Abstract: This study has been made to describe enzymes, especially their kinetics and structures, and is based on the enzyme catalase. The kinetics has been explained by the Michaelis-Menten-equation and
Indledning. MIO er optimeret til Internet Explorer. Læs endvidere under Ofte stillede spørgsmål.
 Indhold Indledning... 3 Søgefunktioner... 4 Søgning fra forsiden... 5 Søgning under menupunktet Instrument... 6 Sådan får man vist instrumenterne i en bestemt afdeling... 7 Sådan ændrer man status på et
Indhold Indledning... 3 Søgefunktioner... 4 Søgning fra forsiden... 5 Søgning under menupunktet Instrument... 6 Sådan får man vist instrumenterne i en bestemt afdeling... 7 Sådan ændrer man status på et
Vejledning til det digitale eksamenssystem. Heilesen, Simon. Publication date: Document Version Peer-review version
 Vejledning til det digitale eksamenssystem Heilesen, Simon Publication date: 2014 Document Version Peer-review version Citation for published version (APA): Heilesen, S. (2014). Vejledning til det digitale
Vejledning til det digitale eksamenssystem Heilesen, Simon Publication date: 2014 Document Version Peer-review version Citation for published version (APA): Heilesen, S. (2014). Vejledning til det digitale
Orcad Layout kørekort. REV. 1.
 Orcad Layout kørekort. REV. 1. Denne guide er skrevet af Carsten Strand fra IHA. Guiden er tænkt som en hjælp til dig der står overfor opgaven at skulle lave et print. Orcad Layout er et omfattende værktøj
Orcad Layout kørekort. REV. 1. Denne guide er skrevet af Carsten Strand fra IHA. Guiden er tænkt som en hjælp til dig der står overfor opgaven at skulle lave et print. Orcad Layout er et omfattende værktøj
Manual til administration af online booking
 2016 Manual til administration af online booking ShopBook Online Med forklaring og eksempler på hvordan man konfigurerer og overvåger online booking. www.obels.dk 1 Introduktion... 4 1.1 Formål... 4 1.2
2016 Manual til administration af online booking ShopBook Online Med forklaring og eksempler på hvordan man konfigurerer og overvåger online booking. www.obels.dk 1 Introduktion... 4 1.1 Formål... 4 1.2
Dannelse af PDF-dokumenter
 Dannelse af PDF-dokumenter Indhold Generere PDF-dokumenter... 2 Håndtering af PDF-dokumentet... 6 Hvordan indsætter man sidetal i PDF-dokumentet?... 6 Hvordan laver man bookmarks i PDF-dokumentet?... 7
Dannelse af PDF-dokumenter Indhold Generere PDF-dokumenter... 2 Håndtering af PDF-dokumentet... 6 Hvordan indsætter man sidetal i PDF-dokumentet?... 6 Hvordan laver man bookmarks i PDF-dokumentet?... 7
ØVELSE 3 ENZYMMÅLINGER OG ENZYMKINETISKE UNDERSØGELSER
 27023, Forår 2013 ØVELSE 3 ENZYMMÅLINGER OG ENZYMKINETISKE UNDERSØGELSER Lærer: Maher Abou Hachem, maha@bio.dtu.dk, Tlf. 45252732, Bygning 224, 122 Indholdsfortegnelse: INTRODUKTION... 3 1. Enzymer - definition
27023, Forår 2013 ØVELSE 3 ENZYMMÅLINGER OG ENZYMKINETISKE UNDERSØGELSER Lærer: Maher Abou Hachem, maha@bio.dtu.dk, Tlf. 45252732, Bygning 224, 122 Indholdsfortegnelse: INTRODUKTION... 3 1. Enzymer - definition
REFWORKS vejledning til Nationale Kliniske Retningslinjer Fagkonsulentens version (december 2013)
") REFWORKS vejledning til Nationale Kliniske Retningslinjer Fagkonsulentens version (december 2013) 2013 Indholdsfortegnelse 1. Refworks og Nationale Kliniske Retningslinjer... 3 2. Log ind i RefWorks...
REFWORKS vejledning til Nationale Kliniske Retningslinjer Fagkonsulentens version (december 2013) 2013 Indholdsfortegnelse 1. Refworks og Nationale Kliniske Retningslinjer... 3 2. Log ind i RefWorks...
WebTV. Vejledning til WebTV på web. Vejledningen beskriver upload og deling af videoer på WebTV
 WebTV Vejledning til WebTV på web Vejledningen beskriver upload og deling af videoer på WebTV ITS 24-11-2015 WebTV Vejledning til WebTV på web Indholdsfortegnelse WebTV... 2 Login... 2 Navigation... 3
WebTV Vejledning til WebTV på web Vejledningen beskriver upload og deling af videoer på WebTV ITS 24-11-2015 WebTV Vejledning til WebTV på web Indholdsfortegnelse WebTV... 2 Login... 2 Navigation... 3
Adobe Acrobat Connect brugergrænsefladen
 Adobe Acrobat Connect brugergrænsefladen Adobe Connect er et webbaseret videokonferenceværktøj, der giver mulighed for online, synkron kommunikation, deling af filer, skærm og whiteboard, gennemførelse
Adobe Acrobat Connect brugergrænsefladen Adobe Connect er et webbaseret videokonferenceværktøj, der giver mulighed for online, synkron kommunikation, deling af filer, skærm og whiteboard, gennemførelse
BentleyUser.dk 2009 MicroStation tips og tricks. Tine Lai Andersen Bentley Institute Instructor
 Tine Lai Andersen Bentley Institute Instructor Brugerflade De små knappenåle / dokkedimser Layouts på dialogbokse Højrekliks menu (laaaangggsomt højreklik): Level Off (V8i SELECT release 1) Display set
Tine Lai Andersen Bentley Institute Instructor Brugerflade De små knappenåle / dokkedimser Layouts på dialogbokse Højrekliks menu (laaaangggsomt højreklik): Level Off (V8i SELECT release 1) Display set
Spm. 1.: Hvis den totale koncentration af monomer betegnes med CT hvad er så sammenhængen mellem CT, [D] og [M]?
![Spm. 1.: Hvis den totale koncentration af monomer betegnes med CT hvad er så sammenhængen mellem CT, [D] og [M]?](/thumbs/54/34467777.jpg "Spm. 1.: Hvis den totale koncentration af monomer betegnes med CT hvad er så sammenhængen mellem CT, [D] og [M]?") DNA-smeltetemperaturbestemmelse KemiF2-2008 DNA-smeltetemperaturbestemmelse Introduktion Oligonucleotider er ofte benyttet til at holde nanopartikler sammen med hinanden. Den ene enkeltstreng er kovalent
DNA-smeltetemperaturbestemmelse KemiF2-2008 DNA-smeltetemperaturbestemmelse Introduktion Oligonucleotider er ofte benyttet til at holde nanopartikler sammen med hinanden. Den ene enkeltstreng er kovalent
Vejledning til indsendelse af artikler via Manuscript Central
 Vejledning til indsendelse af artikler via Manuscript Central Adgang til Manuscript Central o Login o Har du glemt dit password? o Velkomstsiden o Din forfatterside (Author Dashboard) Krav til manuskriptet
Vejledning til indsendelse af artikler via Manuscript Central Adgang til Manuscript Central o Login o Har du glemt dit password? o Velkomstsiden o Din forfatterside (Author Dashboard) Krav til manuskriptet
NVivo-øvelser for PC. Når NVivo er åbent, kan importen ske på to måder:
 NVivo-øvelser for PC Før du går i gang med øvelserne, er det selvsagt nødvendigt at importere øvelsesmaterialet ind i NVivo. Der er her tale om fire nytårstaler (fra 1994, 2002, 2010 og 2012) som Word-dokumenter.
NVivo-øvelser for PC Før du går i gang med øvelserne, er det selvsagt nødvendigt at importere øvelsesmaterialet ind i NVivo. Der er her tale om fire nytårstaler (fra 1994, 2002, 2010 og 2012) som Word-dokumenter.
Kompendium til Geogebra
 Kompendium til Geogebra Hardsyssel Efterskole Matematik 8. Klasse Side 1 af 12 Kompendium til Geogebra 1. Generel præsentation af Geogebra 1.1 Download af programmet Geogebra kan gratis downloades fra
Kompendium til Geogebra Hardsyssel Efterskole Matematik 8. Klasse Side 1 af 12 Kompendium til Geogebra 1. Generel præsentation af Geogebra 1.1 Download af programmet Geogebra kan gratis downloades fra
Start af nyt schematic projekt i Quartus II
 Start af nyt schematic projekt i Quartus II Det følgende er ikke fremstillet som en brugsanvisning der gennemgår alle de muligheder der er omkring oprettelse af et Schematic projekt i Quartus II men kun
Start af nyt schematic projekt i Quartus II Det følgende er ikke fremstillet som en brugsanvisning der gennemgår alle de muligheder der er omkring oprettelse af et Schematic projekt i Quartus II men kun
Portal Registration. Check Junk Mail for activation . 1 Click the hyperlink to take you back to the portal to confirm your registration
 Portal Registration Step 1 Provide the necessary information to create your user. Note: First Name, Last Name and Email have to match exactly to your profile in the Membership system. Step 2 Click on the
Portal Registration Step 1 Provide the necessary information to create your user. Note: First Name, Last Name and Email have to match exactly to your profile in the Membership system. Step 2 Click on the
Sådan opretter du en blog i e-learn.sdu.dk I denne vejledning vises først, hvor i e-learn.sdu.dk, du kan oprette blogs. Derefter vises hvordan du gør.
 Sådan opretter du en blog i e-learn.sdu.dk I denne vejledning vises først, hvor i e-learn.sdu.dk, du kan oprette blogs. Derefter vises hvordan du gør. Du kan oprette blogs på 3 forskellige niveauer i e-learn.sdu.dk:
Sådan opretter du en blog i e-learn.sdu.dk I denne vejledning vises først, hvor i e-learn.sdu.dk, du kan oprette blogs. Derefter vises hvordan du gør. Du kan oprette blogs på 3 forskellige niveauer i e-learn.sdu.dk:
Enzymer og katalysatorer
 Enzymer og katalysatorer Reaktionsligningen: viser den kemiske reaktion, der leverer energi til alle stofskifteprocesser i cellerne i kroppen. Kemisk er der tale om en forbrændingsproces, hvori atmosfærisk
Enzymer og katalysatorer Reaktionsligningen: viser den kemiske reaktion, der leverer energi til alle stofskifteprocesser i cellerne i kroppen. Kemisk er der tale om en forbrændingsproces, hvori atmosfærisk
Quick guide Dynamicweb 9. Kom godt i gang med brugen af redigeringsværktøjet bag vores hjemmesideløsning CMS-systemet Dynamicweb
 Quick guide Dynamicweb 9 Kom godt i gang med brugen af redigeringsværktøjet bag vores hjemmesideløsning CMS-systemet Dynamicweb Indholdsfortegnelse Sådan logger du på... 3 Opbygning... 4 Beskrivelse af
Quick guide Dynamicweb 9 Kom godt i gang med brugen af redigeringsværktøjet bag vores hjemmesideløsning CMS-systemet Dynamicweb Indholdsfortegnelse Sådan logger du på... 3 Opbygning... 4 Beskrivelse af
Gymnasieøvelse i Skanning Tunnel Mikroskopi (STM)
") Gymnasieøvelse i Skanning Tunnel Mikroskopi (STM) Institut for Fysik og Astronomi Aarhus Universitet, Sep 2006. Lars Petersen og Erik Lægsgaard Indledning Denne note skal tjene som en kort introduktion
Gymnasieøvelse i Skanning Tunnel Mikroskopi (STM) Institut for Fysik og Astronomi Aarhus Universitet, Sep 2006. Lars Petersen og Erik Lægsgaard Indledning Denne note skal tjene som en kort introduktion
Help / Hjælp
 Home page Lisa & Petur www.lisapetur.dk Help / Hjælp Help / Hjælp General The purpose of our Homepage is to allow external access to pictures and videos taken/made by the Gunnarsson family. The Association
Home page Lisa & Petur www.lisapetur.dk Help / Hjælp Help / Hjælp General The purpose of our Homepage is to allow external access to pictures and videos taken/made by the Gunnarsson family. The Association
Workshop G4 MicroStation V8 XM edtion nye værktøjer 2
 Workshop G4 MicroStation V8 XM edtion nye værktøjer 2 FØR I BEGYNDER......1 ACCUDRAW...2 ROTATE VIEW...2 ROTATE ELEMENT...2 ANDRE NYE ACCUDRAW SHORTCUTS...2 SHORTCUTS...3 DE VIGTIGSTE...3 CTRL OG EN FUNKTIONSTAST
Workshop G4 MicroStation V8 XM edtion nye værktøjer 2 FØR I BEGYNDER......1 ACCUDRAW...2 ROTATE VIEW...2 ROTATE ELEMENT...2 ANDRE NYE ACCUDRAW SHORTCUTS...2 SHORTCUTS...3 DE VIGTIGSTE...3 CTRL OG EN FUNKTIONSTAST
Fable Kom godt i gang
 Fable Kom godt i gang Opdateret: 26-03-2018 Indholdsfortegnelse 1. Først skal du installere programmet på din computer 3 2. Når programmet er installeret er du klar til at pakke robotten ud 4 3. Nu er
Fable Kom godt i gang Opdateret: 26-03-2018 Indholdsfortegnelse 1. Først skal du installere programmet på din computer 3 2. Når programmet er installeret er du klar til at pakke robotten ud 4 3. Nu er
Jernindhold i fødevarer bestemt ved spektrofotometri
 Bioteknologi 4, Tema 8 Forsøg www.nucleus.dk Linkadresserne fungerer pr. 1.7.2011. Forlaget tager forbehold for evt. ændringer i adresserne. Jernindhold i fødevarer bestemt ved spektrofotometri Formål
Bioteknologi 4, Tema 8 Forsøg www.nucleus.dk Linkadresserne fungerer pr. 1.7.2011. Forlaget tager forbehold for evt. ændringer i adresserne. Jernindhold i fødevarer bestemt ved spektrofotometri Formål